The Low-Cost Engine: Threshold Performance and Recovery in a Long-Term Chen Taijiquan Practitioner
- Tai Chi Gringo
- Mar 2
- 8 min read
Updated: May 20
An observational account of autonomic data that conventional exercise physiology doesn't predict
In the Economics of Effort pillar article, it was established that athletes are conditioned to think in outputs: speed, power, heart rate, rounds completed, minutes spent in Zone 4. Conditioning culture is built around what you can produce. Far less attention is paid to what it costs the system to produce that output, metabolically, neurologically, and systemically.
This article examines whether that picture is complete. Specifically, whether long-term Chen Tai Chi training alters the cost structure of high-intensity work, in the domains of recovery, inflammation, and autonomic regulation, in ways that output-focused models have no framework to account for.
What follows is an observational account of a single subject with an extensive training history in Chen Taijiquan, examined during sustained upper-threshold sparring rounds performed consecutively in tropical heat and humidity. The headline finding: resting heart rate the morning after ten consecutive threshold rounds did not rise. It dropped. HRV did not suppress. It climbed above baseline within 48 hours. This recovery pattern was observed consistently across multiple independent sessions.
The aim of this article is to document the recovery dynamics that follow high-intensity threshold work in this subject, establish that the pattern is reproducible across sessions, and assess whether conventional explanations account for it. The mechanistic question, what produces this pattern, is examined in the companion article on autonomic and metabolic efficiency.
The Training Context
At 42, I had recently begun accumulating sustained threshold work in sparring contexts, kickboxing and boxing, with no history of sustained threshold work in either sport. Occasional sparring rounds across three years of MMA, but nothing approaching the consecutive high-intensity structure of the sessions described here.
My measured cardiac maximum is 212 bpm, substantially above the age-predicted figure of around 178. A recent CPET produced a VO₂ max of approximately 48 ml/kg/min despite no history of specific endurance or threshold training. The session logged 32 minutes between 161 and 179 bpm, a CPET conducted two months later placed VT2 at 172 bpm in a detrained state, anchoring this as genuine threshold work. The stability of the trace across seven consecutive rounds, peaks holding within a 5 bpm band with no progressive drift, confirms a system operating at a high but sustainable ceiling.
The session was conducted in tropical heat and humidity, and was followed immediately by approximately twenty minutes of MMA sparring for which heart rate data was not captured. Total training load that day was considerably higher than the documented session alone represents.
The efficiency pattern this session documents had already been expressing itself across four years of BJJ and MMA training, in recovery dynamics and metabolic examined in BJJ Without Glycolytic Debt, and in structural resilience under sustained grappling demand, examined in Ten Years of Chen Taijiquan Meets High Volume Grappling. The sparring session documented here is the most precisely measured instance of it.
The primary finding of this session is not what happened during it. It is what the body showed following it.




The Recovery Signal
The recovery signal that followed was notable for its absence. Subjective recovery occurred within one to three hours, with no residual heaviness, soreness, or autonomic hangover. Objective data from wearable tracking confirmed this picture across the following days.
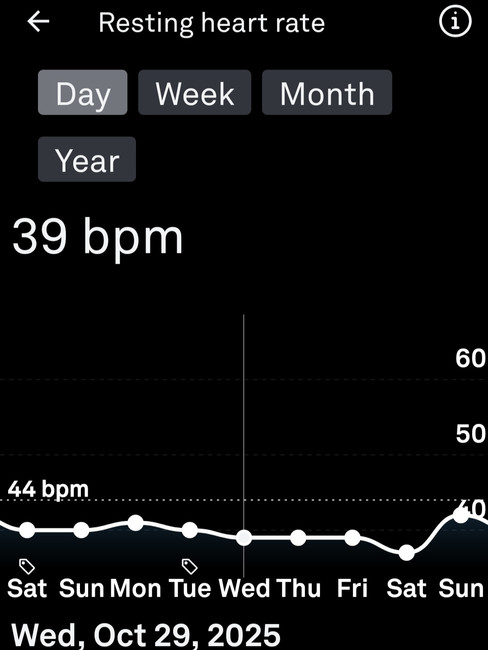
The clearest single finding in the post-session data is the resting heart rate. Pre-session baseline was 42-44 bpm. The first post-session reading on Sunday morning was 43 bpm, squarely within the normal baseline range, as though the session had not occurred. By Sunday night RHR averaged 42 with a low of 39, below baseline. Monday night continued similarly at 41 average and 39 low. By Tuesday night the system had returned to its normal range. There was no elevation arc, no inflammation tail, no autonomic hangover dragging through the week. The data did not just fail to show the expected cost, it showed the opposite of it.
The HRV picture tells the same story through a more sensitive instrument. In the days preceding the session a visiting friend had disrupted normal routine, travel, irregular meals, later nights including New Year's Eve, some alcohol, and HRV reflected this, varying between 40 and 54. The overnight trace following the session shows a genuine suppression signal in the first half of the night, HRV sitting in the low-to-mid 40s as the system managed the acute aftermath of threshold work. What is unusual is what happened next. Rather than remaining suppressed across the full sleep period, as is typical after threshold work, HRV climbed progressively through the second half of the night, peaking at 79 ms, well above the pre-session range. By morning the above-baseline rebound was already established. The suppression arc existed. It simply completed within hours rather than days.
A reasonable skeptic might attribute this to individual variation, a good day, or simply an easy session. That explanation does not survive basic exercise physiology. A genuine threshold session of this structure, 32 minutes in zone 4, ten consecutive rounds, in tropical conditions, should produce measurable next-day consequences in virtually any athlete, and particularly in a 42-year-old. Elevated RHR persisting for 24-48 hours is the standard autonomic stress signature after zone 4 work. HRV suppression is well-documented after threshold sessions and is often more pronounced in trained athletes precisely because their systems are finely calibrated to load. What appears here moves in the opposite direction.




The HRV rebound and RHR suppression documented here are not fitness markers. They are autonomic regulation markers, measuring not how much the system can do, but how rapidly and completely it returns to baseline after genuine high-intensity demand. High cardiovascular fitness does improve some autonomic markers, but trained athletes still show HRV suppression and elevated resting heart rate after hard sessions. That is the expected pattern regardless of fitness level. The divergence here is what fitness alone cannot account for.
The suppression signal in the first half of the night confirms the session generated a genuine autonomic cost. This was not simply an easy session that left no mark. What is unusual is the speed and completeness of its resolution: clearance within hours, followed by a rebound above pre-session baseline before morning. This points to two mechanisms likely operating together. The session probably cost less than the external load would predict, the debt was smaller than expected. And the debt that was generated was cleared faster than normal recovery physiology would suggest.




This is not the signature of a system repaying a recovery debt. It is the signature of a system positively regulated by the effort, left in a measurably better autonomic state than it started. That pattern, suppression resolving within hours, followed by a sustained multi-night rebound below baseline RHR and above baseline HRV, is almost exclusively associated with low-intensity recovery work. Finding it after confirmed threshold sparring in a combat sports context suggests the session functioned not as a sustained stressor but as a regulatory stimulus: a categorically different relationship between high-intensity work and the autonomic system than conventional exercise physiology predicts.
A Consistent Pattern
The recovery pattern documented in the primary session was not an isolated finding. Two further threshold sessions on the 22nd and 25th November, 30 minutes and 24 minutes of zone 4 work respectively, produced the same fundamental response. The second of these occurred just three days after the first, a compressed timeframe that would typically compound recovery debt across sessions. The overnight traces from both sessions are presented below.








The 25th November session shows the most pronounced stress signal across all three documented instances, a modest within-night suppression visible in both heart rate and HRV traces, which is also the session most plausibly carrying residual load from the preceding effort.
Even this clearest signal follows the same pattern: suppression in the first half of the night, resolution and rebound in the second, cleared within a single sleep cycle rather than persisting across days. In both cases the nights following the sessions show HRV climbing above baseline and RHR sitting at or below baseline, the same post-session upregulation documented in detail in the primary session, suggesting the threshold work is functioning as a regulatory stimulus rather than a recovery debt.








A Directly Measured Metabolic Reference Point
The sparring data establishes the recovery pattern through heart rate and wearable autonomic markers alone. The metabolic load of those sessions can only be inferred from the intensity data rather than directly measured. A separate dataset provides the first directly measured metabolic reference point.

During a multi-stage incremental treadmill lactate test performed on 28th October, blood lactate reached 13.9 mmol/L at a heart rate of 170 bpm across the final stage. The overnight Oura data from that night showed a lowest heart rate of 39 bpm, an average overnight heart rate of 44 bpm, an average HRV of 62 ms, and a peak HRV of 95 ms. No meaningful suppression. No elevation of resting heart rate. Autonomic function the same night as a confirmed severe lactate load was essentially indistinguishable from a normal baseline night. In the days following, HRV continued to climb above the pre-test baseline, the same post-stimulus upregulation documented across the sparring sessions.






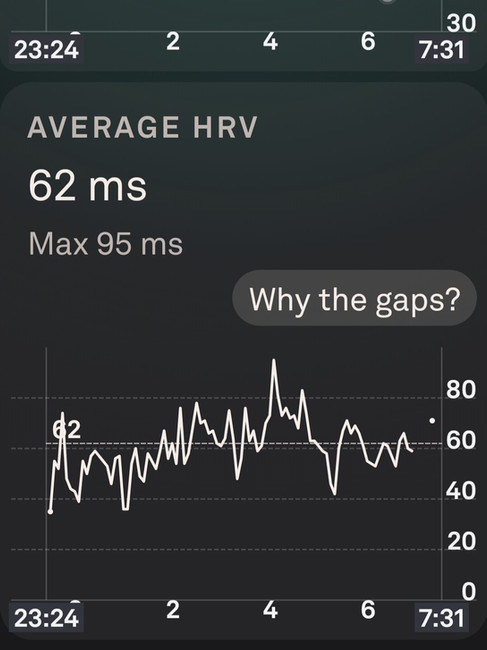
A caveat applies: the treadmill is not an adapted movement pattern, and local muscular fatigue in unadapted running fibres almost certainly contributed to the lactate reading. But 13.9 mmol/L is a systemic measurement. By the time lactate is measurable at that concentration in fingertip blood, it has entered systemic circulation regardless of where it was primarily produced. The buffering systems, hepatic clearance pathways, and catecholamine responses that high systemic lactate triggers are activated by the circulating lactate itself, not its origin. The conventional prediction for overnight autonomic data following that exposure is suppressed HRV and elevated resting heart rate.
What the overnight data showed was the opposite. Peak HRV of 95 ms. Resting heart rate at 39 bpm. These are not the numbers of a system managing a severe metabolic debt. They are the numbers of a system in strong parasympathetic dominance, the same pattern the sparring data documents, now grounded in a directly measured metabolic load rather than an inferred one.
It is worth clarifying what high lactate actually predicts. Lactate is a fuel and signalling molecule, modern exercise physiology is clear that it is not itself the primary driver of recovery cost. The real systemic burden comes from muscle damage, glycogen depletion, central fatigue, and sympathetic activation, all of which can decouple substantially from lactate levels. The question the overnight data raises is therefore not why 13.9 mmol/L produced low recovery cost, it is why the session produced high lactate without the muscle damage and sympathetic activation that conventionally accompany it. The mechanisms that may account for that decoupling are examined in companion pieces: the autonomic and metabolic layer in the piece on proposed mechanisms driving the low cost engine adaptation, the structural and neurological layer in the piece on internal training and systemic efficiency.
What the Data Can and Cannot Establish
The sub-baseline RHR and above-baseline HRV rebound after confirmed threshold sparring in Bali's tropical conditions is genuinely unusual, not just for the general population but for trained athletes, and more so in a combat sports context than a steady-state endurance one. That it occurred consistently in a 42-year-old makes it more striking again; recovery capacity declines with age in ways that would predict a larger suppression signal, not a smaller one. That is the finding, stated with confidence. What produced it remains a question of inference rather than proof.
The most interesting adaptations are often invisible until the system is stressed. Sustained threshold sparring did not reveal a fragile body pushed past its limits. It revealed a regulated system, a 42-year-old chassis operating at younger RPMs with surprising economy.
If internal training genuinely reduces the metabolic debt of hard sessions and compresses the recovery window, and the data presented here suggests it does, the consequences extend well beyond any single workout. Training volume is one of the most robust predictors of athletic development across virtually every discipline. The athlete who can sustain five hard sessions a week instead of three accumulates a dramatically different stimulus over months and years. For the aging athlete, whose recovery capacity is already declining in ways that progressively constrain sustainable volume, that compression is sharpest, an offset against a trajectory that conventional training cannot reverse.
Internal martial arts don't raise the redline. They quietly lower the cost of operating near it. And that difference only becomes visible when the system is finally stressed hard enough to reveal it.



Comments